


ホオジロザメもシュモクザメもジンベエザメも、私たちはふつう、まとめて「サメ」と呼びます。
海を切るように泳ぎ、鋭い歯を持ち、見た目にもいかにも同じ仲間だからです。
けれど今回の研究は、その当たり前に静かな揺さぶりをかけました。
アメリカのイェール大学(Yale)で行われた研究ではサメやエイ、ガンギエイ、ギンザメの仲間を含む48種のゲノム規模の情報が詳しく調べられ、「サメ」は進化の上でひとまとまりのグループではない可能性が示されました。
さらに驚くべきことに、非常に変わりにくいDNAの部分を使った解析では、私たちがふだん「いわゆるサメ」と思っている有名な種の多くは、古いタイプのサメよりも、むしろエイの仲間に近い可能性があると示されました。
つまり「サメ」という言葉は、海での暮らしぶりをうまくまとめた便利な呼び名ではあっても、ウマやネコのように、進化の木の枝をそのまま切り出した名前ではないかもしれません。
では、私たちがサメだと信じてきた生き物たちは、本当はどのような歴史の分かれ道をたどってきたのでしょうか。
サメらしさとは何なのか、今回の研究はその核に迫ります。
研究内容の詳細は2026年2月15日にプレプリントサーバーである『bioRxiv』にて発表されました。
目次
- 「サメ」という箱は進化的に幻想だった
- 「超保存領域」が暴いたサメの過去
- サメたちは恐竜時代の「新参者」だったかもしれない
「サメ」という箱は進化的に幻想だった

家系図で最も重要なのは、いちばん最近どこで枝分かれしたかです。
中学生で習う自分との近さを表す一親等、二親等、三親等…という表現も、枝分かれ部分が重要になってきます。
生物の進化系統樹も同じであり、2つの種の近さを比べる時も、2つの種の枝分かれがいつ起きたかや、枝と枝がどれだけ離れているかより、どこで枝分かれしたかが分類において重要になります。
そして私たちは枝分かれ情報をもとに、一定の範囲を囲んで、「ウマ」や「ネコ」など、おおくくりな名前をつけます。
ところが新たな研究では、わたしたちが「いわゆるサメ」と呼んでいる存在に対して、大きな疑問が突き付けられました。
今回の研究では、サメやエイの仲間四十八種のゲノム規模の情報を使って、その家系図がくわしく調べられました。
すると、ふつうは同じ箱に入っている「サメ」が、進化の木の上ではきれいなひとかたまりではない可能性が示されたのです。
たとえば
①かなりマイナーな超古い時代から分かれた、もはやサメではない近縁種(ギンザメ類)
②あまりメジャーではない古い時代からいるサメ(六鰓目)
③よくみるメジャーなタイプの「いわゆるサメ」(ホオジロザメなど)
④実はサメに近い生物として知られるエイ類
という区分があったとき、私たちは②と③に対して「サメ」、④は「エイ」、①は「ギンザメ」とおおくくりな名前をつけます。
しかし新たな研究では、非常に変わりにくいDNAの部分を使った解析では、③の「いわゆるサメ」に一番近いのが④のエイ類たちであり、次いで②の古いサメ、①のギンザメ類という順番である可能性が示されました。
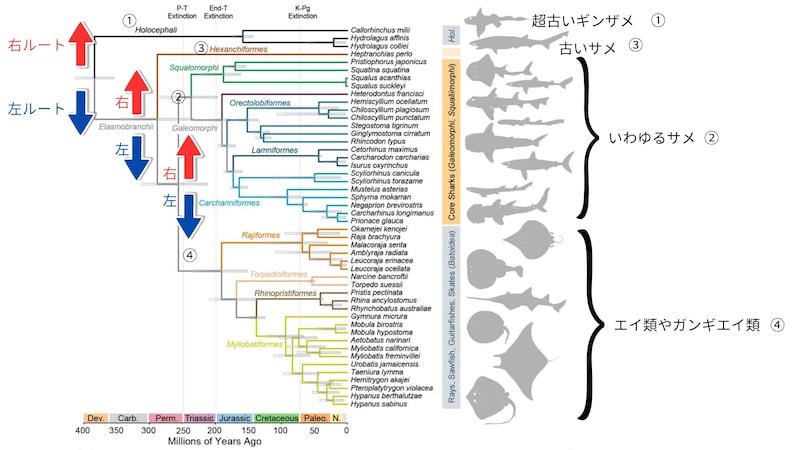
具体的にはまず最初の大きな分かれ道で、右に進んだのがつまりギンザメ類(Holocephali)のルートで、左に進んだのが、サメ・エイをまとめた区分(Elasmobranchii)です。
共通の大きな祖先からギンザメ類は早くから分岐したことから、ギンザメ類はサメやエイではなく、別の近縁のグループという認識になっています。
そして、左ルートを進んだサメ・エイをまとめた区分(Elasmobranchii)はさらに右ルートで六鰓目という「古いサメ」と左ルートでそれ以外にわかれました。
問題はここからです。
そこからさらに左ルートを進んだものたちが「いわゆるサメ」と「エイの仲間」に別れました。
その結果「いわゆるサメ」に一番近い位置に「エイの仲間」が来て、二番目に「古いサメ(六鰓目)」が置かれ、ギンザメ類はそのさらに外側にいるという奇妙な事態が見えてきたのです。

これを家系と財産譲渡で考えるとわかりやすいと思います。
たとえば、鮫家の鮫太郎さんはこれまでずっと、自分に一番近いのは同じ家に住む同じ鮫姓を持つ叔父の鮫兵衛さんだと思い、財産を譲ってきました。
鮫太郎さんの中では「叔父の鮫兵衛さんこそ鮫家の仲間」という認識があったからです。
ところが、突然新たな系譜が発見され、実は英(エイ)の姓を持った英三郎さんが鮫太郎さんの兄弟だったとわかったのです。
すると近い血筋を優先して財産を譲るならば、直近で分岐した兄弟にあたる英三郎が最優先になります。
「なにを当然なことを言ってるんだ」「叔父と兄弟なら兄弟が優先なのはあたりまえ」と思われるかもしれませんが、重要なのは、これまで信じられていた「鮫家という区分」が間違っていたという点です。
同様の話を今回の話に当てはめると、「いわゆるサメ」の兄弟の位置に「エイ類」がいて、叔父の位置に「サメ(古い)」がいる状態です。
そうなると、これまでなんとなく使ってきた「サメ」という言葉は、見た目をうまくまとめた日常語ではあっても、進化の木の枝という観点からみれば、ひと固まりに囲えるという考えは幻想となっていきます。
もちろん、海で背びれのある細長い影を見れば、これからも私たちはそれをサメと呼ぶはずです。
その呼び方が急になくなるわけではありません。
ですが科学が見ているのは、私たちの直感ではなく、はるか昔の枝分かれの順番です。
その目で見たとき、「サメ」という当たり前の言葉が進化の枝分かれの実際と一致していない可能性があるのです。
「超保存領域」が暴いたサメの過去

では、なぜこんな大きな話になったのでしょうか。
通常、進化の枝分かれを調べる一つの方法として、DNAのタンパク質の設計図部分の違いを比較します。
しかし新たな研究では、非常に変化しにくい「超保存領域」と呼ばれる部分も比較されました。
超保存領域とは、長い長い進化の時間をへても、ほとんど姿を変えずに残ってきたゲノムの部分のことです。
タンパク質の設計図部分にくらべて、ここの変化は非常にゆっくりです。
では、なぜそんな部分が家系図づくりに役立つのでしょうか。
理由は、あまりに変わりやすい場所ばかりを見ると、古い昔の枝分かれをたどるのが難しくなることがあるからです。
何億年も前の分かれ道を知りたいときには、短い時間でどんどん変わる部分より、ゆっくり変わる部分のほうが手がかりになりやすいのです。
するとタンパク質の設計図部分をもとにしたデータからは、サメは今まで通り、進化の枝の上でもひとまとまりの「いわゆるサメ」のグループとして分類できました。
ところが超保存領域の解析では、六鰓目と呼ばれる「古いサメ」が「いわゆるサメ」から飛び出し、こちらでは早い時期に分岐していることがわかったのです。
今回の研究では、今生きている軟骨魚類の共通祖先はおよそ三億八千万年前、六鰓目が他のサメとエイから分かれたのはおよそ二億八千万年前、そしてそれ以外のサメとエイの共通祖先はおよそ二億五千万年前と見積もられています。
研究者たちはこのような億年単位の進化系統樹を調べるには、超保存領域のデータのほうがやや有力になり得ると考えているようです。
もしこの予測が正しければ、私たちが「いわゆるサメ」と呼んでいるグループでは、主要な枝分かれが恐竜時代の前半から中ごろにかけて集中したとされます。